豆渣固态发酵过程中主要营养成分及抗氧化特性变化

导读
大豆是亚洲地区重要的粮油作物之一。随着豆制品加工技术的成熟和消费需求的增长,作为豆浆、豆腐或其他大豆产品生产中所产生的主要副产品,豆渣除小部分作为牲畜饲料外,大部分被当成工业废料丢弃。然而,豆渣含有丰富的膳食纤维、蛋白质、脂肪、异黄酮和维生素等,具有丰富的营养价值。目前,世界上越来越多的国家将豆渣看作是一种新的保健食品源,在我国豆渣资源尚未被充分利用。
固态发酵技术是一种经济方便的发酵技术,被广泛地应用于食品生产和加工过程中。微生物在发酵过程中所产生的复杂酶系和众多活性物质,可以提高豆渣的营养价值、改善口感、提高产品的生物活性。近几年来,使用豆渣进行固态发酵生产一些具有生物活性的化合物的研究日益增多,李红艳等报道豆渣经粗壮脉纹孢菌发酵,类胡萝卜素含量大大增加。Zhu Yunping等利用枯草杆菌(Bacillus subtilisB2)固态发酵,可以得到一种叫做1-脱氧野尻霉素(1-deoxynojirimycin,DNJ)的物质,DNJ是一种糖苷酶抑制剂,可以用于糖尿病患者。还有研究表明,可以利用青霉菌发酵豆渣得到新型杀虫剂。但国内将豆渣直接应用开发为功能性食品的研究以及对发酵豆渣食品特性的研究较少。
在国外,Kronenberg、Matuso等对微生物发酵豆渣的营养成分及食品特性进行了研究,而均未对发酵豆渣的抗氧化功能特性进行深入研究。为此本实验以传统固态发酵工艺为基础,研究了米根霉及少孢根霉两株霉菌在豆渣发酵过程的情况,对发酵过程中主要营养成分及抗氧化活性进行测定,与未发酵的豆渣进行对比,为开发利用豆渣制成的功能性食品提供理论参考。
1.1 材料、菌株与试剂
大豆购买于江苏省南京市苏果超市。少孢根霉(Rhizopus oligosporus RT-3)、米根霉(Rhizopus oryzae)均由南京农业大学食品微生物实验室保存。
2,2’-联氮-二(3-乙基-苯并噻唑-6-磺酸)二铵盐(2,2’-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid),ABTS)、菲洛嗪、水溶性VE(Trolox) 美国Sigma公司;乙二胺四乙酸二钠(ethylenediaminetetraacetic aciddisodium salt,EDTA-2Na)、氯化铁、硫酸亚铁、铁氰化钾、三氯乙酸等均为分析纯试剂。
1.2 仪器与设备
TM2300全自动凯氏定氮仪丹麦Foss公司;微波消解仪意大利Milestone公司;索式脂肪抽提装置上海华瑞仪器有限公司;SX2-4-13数显控温马弗炉上海苏净实业有限公司;LHS-150SC恒温恒湿箱上海一恒科技有限公司;AUY-120分析天平、UV-2450紫外分光光度计日本Shimadzu公司;HH-4数显恒温水浴锅国华电器有限公司;酶标仪 美国Bio-Tek公司;64RL高速冷冻离心机美国Beckeman公司;电热恒温鼓风干燥箱上海益恒实验仪器有限公司;手提式不锈钢蒸汽消毒器上海三申医疗器械有限公司。
1.3 方法
1.3.1 发酵豆渣的制备
称取大豆1 000 g,加入3 000 mL蒸馏水,室温条件下浸泡12 h。以1∶7(m/V)的豆水比进行磨浆,制得含水量85%的新鲜豆渣。将新鲜豆渣烘至含水量为60%后放在121 ℃的高压灭菌锅内灭菌15 min。在其中一份豆渣中按1 g/100 mL的接种量接种米根霉的孢子悬浮液(108~109个/mL),另一份豆渣中按同样的接种量接种少孢根霉的孢子悬浮液(108~109个/mL)。混匀后装入保鲜袋中并铺平。铺平后的豆渣约占保鲜袋面积的1/2,厚度约3 cm,每隔2 cm扎小孔透气后,置于30 ℃培养箱中发酵30 h。每隔6 h取样,将取出的样品冷冻并经粉碎机粉碎,过80 目筛备用。
1.3.2 发酵豆渣理化指标测定
1.3.2.1 膳食纤维含量的测定
参照GB/T 5009.88—2008《食品中膳食纤维的测定》。
1.3.2.2 粗蛋白含量的测定
参照GB 5009.5—2010《食品中蛋白质的测定》。
1.3.2.3 粗脂肪含量的测定
参照GB/T 14772—2008《食品中粗脂肪的测定》。
1.3.2.4 灰分含量的测定
参照GB 5009.4—2010《食品中灰分的测定》。
1.3.2.5 还原糖含量的测定
1 g冻干样品置于25 mL 蒸馏水中,50 ℃水浴摇床提取30 min,4 ℃、8 000×g离心15 min,取上清液定容至100 mL待测。采用3,5-二硝基水杨酸比色法测定样品还原糖含量。以葡萄糖质量浓度为横坐标,吸光度为纵坐标绘制标准曲线。结果以葡萄糖当量表示(mg/g,以豆渣冻干粉计)。
1.3.2.6 可溶性蛋白含量的测定
1 g冻干样品置于25 mL pH 9.0 硼酸-氢氧化钠缓冲液,室温条件下超声辅助提取1 h,4 ℃、8 000×g离心15 min,取上清液定容至25 mL待测。采用考马斯亮蓝法测定样品中的可溶性蛋白含量。以牛血清白蛋白质量浓度为横坐标,吸光度为纵坐标绘制标准曲线。结果以牛血清白蛋白当量表示(mg BSA/g,以豆渣冻干粉计)。
1.3.2.7 纤维素酶活力测定
1 g冻干样品置于20 mL 0.1 mol/L pH 4.8的乙酸钠缓冲溶液中,磁力搅拌30 min,4 ℃、8 000×g离心15 min,上清液则为粗酶液。纤维素酶活力的测定采用文献报道的方法。
酶活力单位(U)定义:50 ℃、pH 4.8时每分钟水解生成1 μg葡萄糖所需要的酶量。
1.3.2.8 糖化酶活力测定
粗酶液制备方法同1.3.2.7节。糖化酶活力的测定采用文献报道的方法。
酶活力单位(U)定义:30 ℃、pH 4.8时每分钟水解生成1 mg葡萄糖所需要的酶量。
1.3.2.9 蛋白酶活力测定
以1∶20的料液比于蒸馏水中室温条件下提取1 h,4 ℃、8 000×g离心15 min,上清液则为粗酶液。参照GB/T 23527—2009《蛋白酶制剂》中关于蛋白酶活力测定的方法测定样品中的蛋白酶活力。
酶活力单位(U)定义:40 ℃,pH 7.5时每分钟从可溶性酪蛋白中水解生成1 μg酪氨酸所需要的酶量。
1.3.3 发酵豆渣抗氧化活性的测定
1.3.3.1 水溶性提取物的制备
1 g冻干样品置于25 mL蒸馏水,50 ℃水浴摇床提取4 h,8 000×g离心15 min,定容至25 mL,上清液以0.45 μm膜过滤,即为发酵豆渣水溶性提取物。
1.3.3.2 还原力测定
还原力的测定采用文献报道的方法。先后在200 μL样品溶液中加入1 mL 0.2 mol/L pH 6.6的磷酸盐缓冲液和1 mL 1 g/100 mL铁氰化钾溶液。混匀,50 ℃水浴20 min,取出后流水迅速冷却,再加入1 mL 10 g/100 mL三氯乙酸。取1 mL上清液加入200 μL 1 mg/mL氯化铁溶液。10 min后于700 nm波长处测吸光度。以Trolox质量浓度为横坐标,吸光度为纵坐标,绘制标准曲线。结果以Trolox的当量表示(μg Trolox/g,以豆渣冻干粉计)。
1.3.3.3 清除ABTS+•活性测定
清除ABTS+·能力的测定按照文献报道的方法。10 mL的7 mmol/LABTS储备液与20 mL 2.45 mmol/LK2S2O8溶液混合后,室温下暗反应16 h,形成ABTS+•。使用前用0.01 mol/L磷酸缓冲液稀释至734 nm波长处吸光度为0.70±0.05时,作为本实验的ABTS工作液。测定时,取1 mL样品溶液与4 mL ABTS+•工作液混合,室温条件下静置反应6 min后,迅速测定其在734 nm波长处的吸光度。以Trolox质量浓度为横坐标,ABTS+•清除率为纵坐标,绘制标准曲线。结果以Trolox的当量表示(μgTrolox/g,以豆渣冻干粉计)。
1.3.3.4 金属离子螯合能力测定
金属离子螯合能力的测定按照文献报道的方法。先后加入0.50 mL样品溶液、0.75 mL蒸馏水、2 mmol/L氯化亚铁溶液,混匀后加入0.10 mL 5 mmol/L菲啰嗪室温反应20 min,测定562 nm波长处吸光度。以EDTA-2Na质量浓度为横坐标,金属离子螯合率为纵坐标,绘制标准曲线。结果以EDTA-2Na的当量表示(μg EDTA-2Na/g,以豆渣冻干粉计)。
1.4 数据处理与分析
实验数据均为3 次重复实验所得的平均值,结果表示为±s。采用SPSS 16.0软件进行显著性和相关性分析,采用Origin 8.5软件作图。
2结果和分析
2.1 发酵豆渣的一般化学成分分析
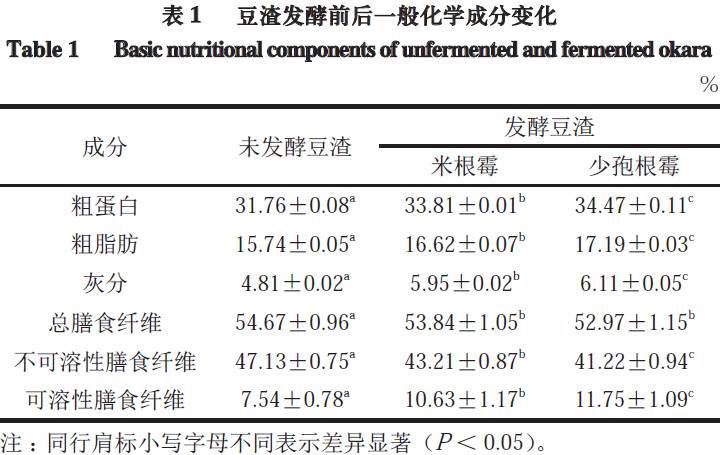
对未发酵豆渣及由米根霉、少孢根霉发酵的豆渣冷冻干燥后,进行粗蛋白、粗脂肪、灰分、膳食纤维含量的测定(表1)。未发酵的豆渣蛋白质含量为31.76%,经过米根霉、少孢根霉发酵后,蛋白质含量分别提高至33.81%、34.47%。同时粗脂肪、灰分的含量也有所提高。一方面由于伴随着微生物的生长,霉菌菌丝覆盖整个豆渣表面,霉菌菌丝与豆渣基质合二为一,菌丝中的粗蛋白、粗脂肪、灰分均能够引起发酵豆渣中相应含量的提高。另一方面发酵过程中随着微生物生长,豆渣中的部分物质被微生物当做碳源利用,导致其他营养成分的比例发生改变,从而导致发酵后其化学成分的含量发生改变。如微生物在发酵过程中将豆渣中的某些营养物质降解为一些小分子物质,如醇和酸发生酯化反应,形成酯类物质,从而使得乙醚提取物增加,导致粗脂肪含量明显增加(P<0.05)。豆渣中总膳食纤维含量很高,占豆渣干质量的54.67%,其中主要成分是不可溶性膳食纤维(insoluble dietary fiber,IDF),含量为47.13%,而可溶性膳食纤维(soluble dietary fiber,SDF)含量相对较低,只有7.54%。发酵后,IDF含量有所下降,SDF的含量有所上升,同时IDF和SDF的比值显著下降(P<0.05)。发酵前IDF/SDF为6.25,经过米根霉、少孢根霉发酵后IDF/SDF分别降低到4.06、3.51,这表明发酵过程中,微生物会产生少量的纤维素酶,将一部分IDF水解成SDF,从而造成IDF/SDF值的变化。
2.2 还原糖含量变化
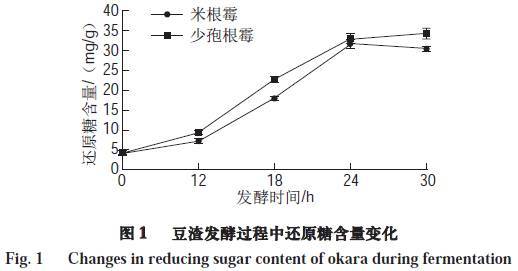
由图1可知,豆渣中的还原糖随着发酵时间的延长而增加,其中在12~24 h期间显著增加(P<0.05),经米根霉、少孢根霉发酵30 h后豆渣还原糖含量分别为30.47、34.28 mg/g,比未发酵豆渣分别提高了7.31、8.22 倍。由于在发酵过程中微生物所分泌的纤维素酶及淀粉酶的作用,可将豆渣中的纤维素、淀粉等降解为低分子的还原糖,从而使豆渣中还原糖的含量随着发酵时间的延长而增加。
2.3 可溶性蛋白含量变化
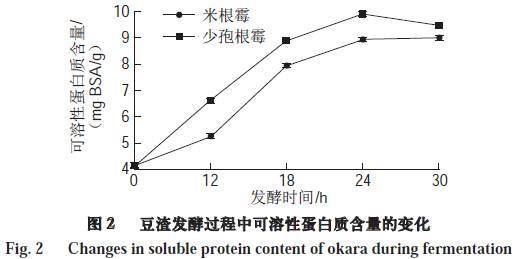
由图2可知,随着发酵时间的延长,豆渣的可溶性蛋白质含量在0~24 h显著增加(P<0.5),之后稍有下降。少孢根霉发酵豆渣的可溶性蛋白质含量由发酵前的4.17 mg BSA /g增加至9.44 mg BSA /g,米根霉发酵豆渣的略低于同时间内少孢根霉发酵豆渣,为8.99 mg BSA/g。由于在微生物的发酵过程中,豆渣中的蛋白质被分解成多肽和一些有一定空间结构但分子质量较小的蛋白质,增加了其蛋白质的溶解性,所以可溶性蛋白质的含量会增加。Sun Hong等采用枯草芽孢杆菌对棉籽粉进行发酵,也发现发酵后的棉籽粉中可溶性蛋白质增多。进一步延长发酵时间,豆渣中的蛋白质更多的被降解成氨基酸和分子质量更小的小肽物质,而考马斯亮蓝法更多是反映大分子可溶性蛋白质变化的情况,无法准确测定其中的氨基酸和小肽,从而造成后期可溶性蛋白质呈现下降趋势。
2.4 纤维素酶活力变化
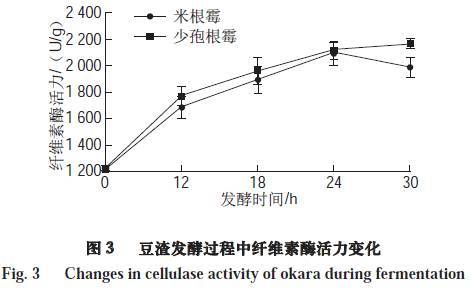
由图3可知,发酵后的豆渣中纤维素酶活力较未发酵的豆渣有显著提高(P<0.05)。发酵30 h后,经米根霉、少孢根霉发酵的豆渣中纤维素酶活力分别达到1 989.42、2 165.73 U/g,比未发酵豆渣分别提高了1.62、1.77 倍。纤维素酶活力的变化影响豆渣中膳食纤维的含量。微生物在发酵过程中所产生的纤维素酶将水不溶性膳食纤维多糖水解,从而使水溶性膳食纤维多糖成分增加。另一方面水不溶性膳食纤维的大大减少导致总膳食纤维含量的降低。霉菌发酵可适当提高豆渣可溶性膳食纤维含量,使得豆渣的抗氧化、抗血压等功能活性提高。
2.5 糖化酶活力变化
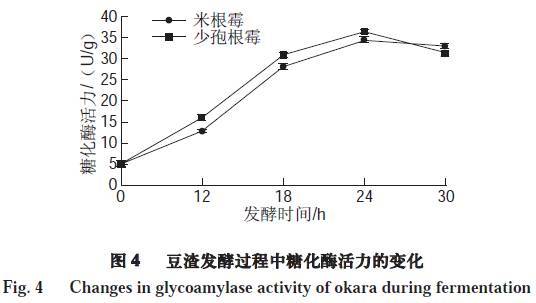
由图4可知,未发酵豆渣中糖化酶活力较低,仅为4.90 U/g,经米根霉、少孢根霉发酵后,其糖化酶活力与未发酵的豆渣相比得到显著提高(P<0.05)。米根霉、少孢根霉发酵的豆渣糖化酶活力均在发酵24 h达到最大,最高值分别为34.25、36.09 U/g。微生物可以在发酵过程中分泌产生糖化酶,将豆渣中的碳水化合物糖化,进而提高微生物生长所需要的碳源。继续培养,糖化酶活力均稍有下降,此时霉菌生长已经处于停滞期,抑制糖化酶活力,从而导致酶活力下降。糖化酶活力的变化与还原糖变化规律基本一致。
2.6 蛋白酶活力变化
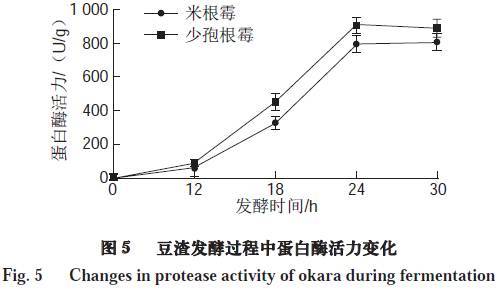
由图5可知,豆渣在初始发酵的12 h内,蛋白酶活力变化不明显,可能是因为在0~12 h霉菌孢子处于萌发状态。12 h后,随着发酵时间的延长,微生物生长迅速,代谢也逐渐旺盛,蛋白酶活力不断提高。在24 h经米根霉、少孢根霉发酵的豆渣中蛋白酶活力分别达到796.68、906.47 U/g。但是,随着发酵时间的进一步延长,微生物分解蛋白质的能力减弱,从而导致蛋白酶活力变化趋于稳定。微生物在发酵过程中所产生的蛋白酶会使得豆渣中的功能性小肽和氨基酸得到部分积累,从而改善豆渣的功能活性。
2.7 发酵豆渣的抗氧化能力
由于不同的自由基和损伤剂会导致不同的氧化损伤,因此单一的抗氧化体系很难全面体现其生物学意义,需要相互补充的多种体系来评价样品在不同体系中的真实效应。本研究采用还原能力、ABTS+·清除能力以及金属离子螯合能力3 种不同的抗氧化能力评价指标,试图从多个侧面反映豆渣在发酵过程中抗氧化能力的变化。
2.7.1 还原力变化
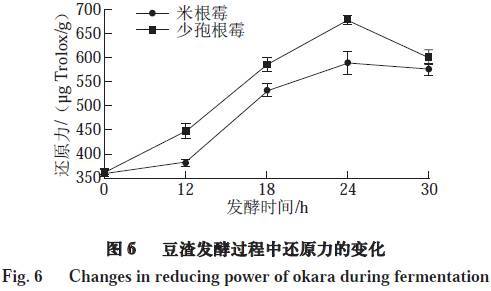
由图6可知,对于少孢根霉和米根霉两株菌而言,其还原力均随着发酵时间的延长而明显增强。且两株菌的还原力均在发酵24 h时达到最大值,分别为677.70、588.24 μg Trolox/g。Yang等认为在发酵过程中产生了一些还原酮类物质,这些物质可通过提供电子使自由基变为稳定的物质,以中断自由基链式反应,从而导致发酵后的样品具有较强的还原能力。除此之外,蛋白质的水解产物如小肽物质和氨基酸,如亮氨酸、蛋氨酸、酪氨酸、组氨酸和色氨酸也能使发酵后的样品具有较强的还原能力。随着发酵的进行,微生物所分泌的各种酶类物质,如纤维素酶、蛋白酶、糖化酶的产生,使发酵后的样品产生更多的还原糖、小肽和氨基酸等物质,这些都与发酵后样品还原能力的提高有关。
2.7.2 ABTS+·清除活性
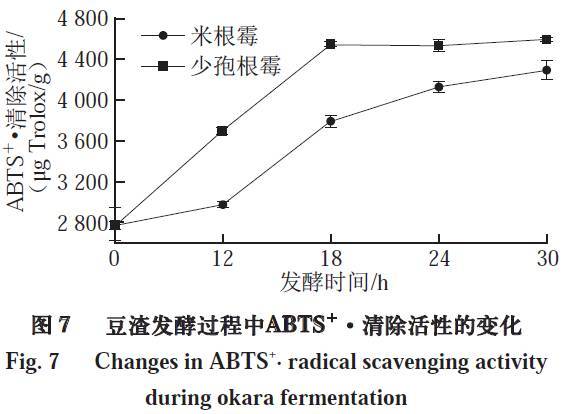
如图7所示,米根霉、少孢根霉在发酵0~18 h时,豆渣的ABTS+·清除活性均随着发酵时间的延长显著上升,在18 h分别达到3 803.34、4 546.31 μg Trolox/g,相比于未发酵的豆渣分别提高了1.36、1.63 倍。少孢根霉发酵豆渣的ABTS+·清除活性在18 h后趋于稳定,而米根霉发酵豆渣的在18 h后随着发酵时间的延长仍有小幅度提高。在30 h时,其ABTS+·清除活性为4 298.66 μg Trolox/g。ABTS+·清除能力的不同表明发酵产物的抗氧化性与其抗氧化物的种类和含量有关。不同微生物有不同的发酵特性,因此在发酵过程中,发酵产物的种类、比例以及含量都会随着发酵时间的进行而变化,因此捕获自由基的能力可能有变化。
2.7.3 金属离子螯合能力
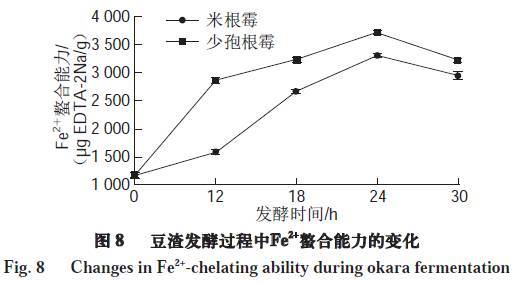
如图8所示,米根霉和少孢根霉发酵24 h后其水提取物对于Fe2+螯合能力比未发酵豆渣有明显提高,螯合能力分别为3 296.88、3 709.16 μg EDTA-2Na/g,此后随着发酵时间的延长稍有降低。已有报道显示许多传统发酵食品,如koji、kinema在发酵后均能提高其对金属离子的螯合能力。Pownall等指出一些功能性小肽由于其特殊的肽链结构及侧链氨基酸,可以阻断自由基的链式反应,对于螯合过渡态的金属离子起着重要的作用。
3讨论
用米根霉、少孢根霉为发酵菌株,以鲜豆渣为原料,纯种发酵制备发酵豆渣。发现在发酵过程中,随着发酵时间的延长会引起发酵基质产生大量的生物化学变化,主要体现在还原糖含量、可溶性蛋白质含量、纤维素酶活力、糖化酶活力和蛋白酶活力都随着发酵时间的延长显著增加,也说明了在发酵过程中米根霉和少孢根霉能够在豆渣中较好的生长,将许多大分子物质有效分解为易被吸收利用的小分子物质,为微生物的后期发酵提供充足的碳源、氮源。两种菌株发酵制得的豆渣其各自的水提取物都表现出较强的还原能力、ABTS+·清除能力、Fe2+螯合能力,说明发酵豆渣与未发酵豆渣相比,具有更强的抗氧化活性,可以作为氢供体、自由基清除剂以及过氧化金属离子的螯合剂,为豆渣的功能性食品开发提供了可能性。
- 上一篇:贵港正邦农牧科技有限公司举办规模猪场互联大会,整合广西助农等资源,开创新篇章 [2017-11-24]
- 下一篇:防城港市举办两期广西现代生态养殖猪和牛的培训班,特邀广西助农公司技术员专业培训 [2017-11-23]






